| Lecture
#36 |
| Hemoglobin...an
accumulation of all the chemistry we have seen in this
semester course. |
CURMUDGEON
GENERAL'S WARNING. These "slides" represent
highlights from lecture and are neither complete nor
meant to replace lecture. It is advised not
to use these as a reliable means to replace missed
lecture material. Do so at risk to healthy academic
performance in 09-105. |
| Lecture Outline |
Hemoglobin
Bonding of heme Fe2+ to O2
vs CO
Sizes of transition metal ions
Role of protein in hemoglobin
|
| Amino acids are oxyacids in which the -OH group is
bound to a carbon. That carbon is also double-bonded to a
=O and single-bonded to an "amino"
(ammonia-like) -NH2 group.Amino acids are
connected by peptide bonds to form polypeptides. As soon
as you have a number of amino acids joined, the
combinatoric possibilities become staggering. |
 |
| How many different arrangements of 300 amino acids
are possible? |
 |
| The number of different possibilities is
large...indeed! The universe isn't big enough to be
packed (I mean packed) with just one of
each kind. |
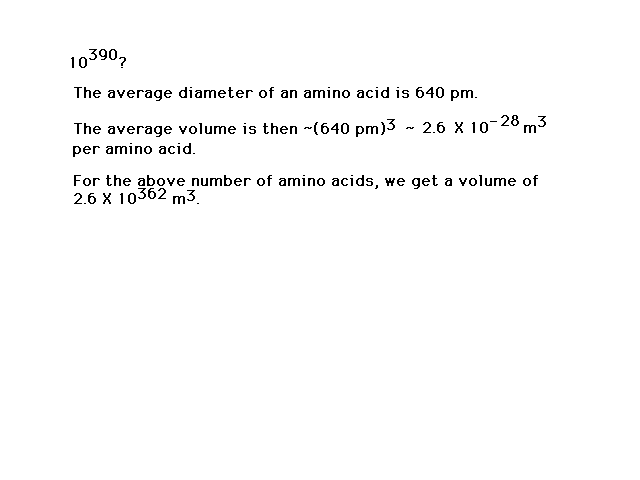 |
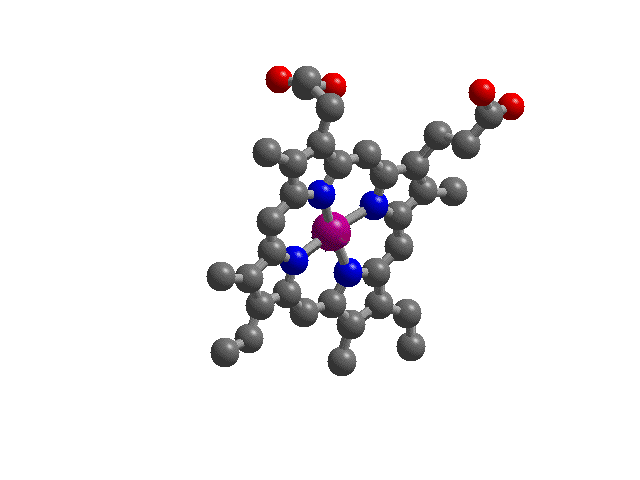
| Hemoglobin: "Heme",
the complex iron ion molecule that runs the essential
chemistry of oxygen transport by blood. In blood, four
hemes are part of a large protein structure called
"hemoglobin". Shown here is "heme"
with the divalent iron ion in purple at its center and
the four (blue) nitrogen donor atoms as part of the heme
ring coordinating ligand.
|
 |
| The chemistry of oxygen in hemoglobin starts with a
look at the Fe2+ at the heme site. |
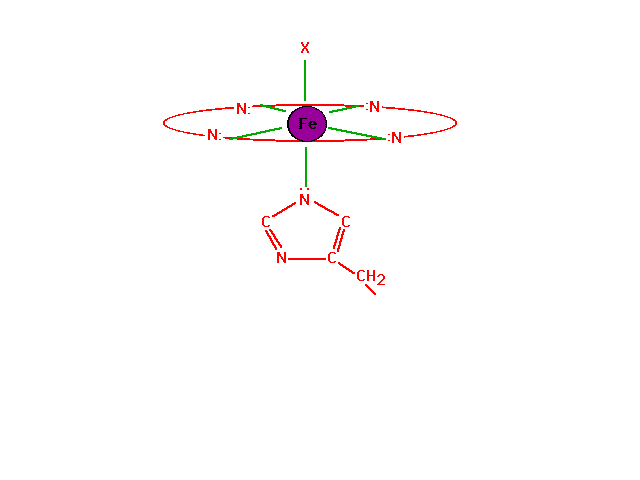 |
| The chemistry of oxygen in hemoglobin starts with a
look at the Fe2+ at the heme site. |
|
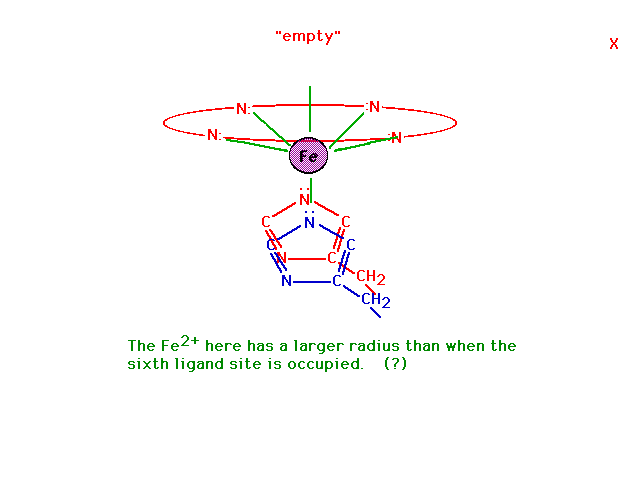
| In the absence of oxygen at the sixth octahedral
site, the geometry of the complex ion changes and this
geometric shift is mechanically transmitted to the other
heme sites on the hemoglobin molecule making it easier
for those to give up their oxygens as well. This leads to
cooperativity among sites for both oxygen release (at
cells being fed oxygen) and also oxygen takeup (by red
blood cells in the lungs). Why the geometry changes has
to do with the ability of a small ion to fit in the
planar coordination site and a large ion to be pushed
out. |
 |
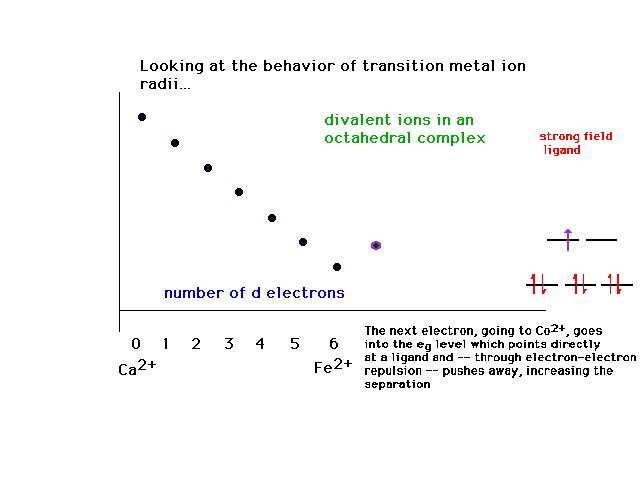
| The effects of adding electrons across the transition
metal ion series produces some surprises, although
they're understandable, showing the influence of the
complexes' geometries and ligand character. This is the
behavior of ionic radii within a complex ion for strong
field octahedral ligands. |
 |
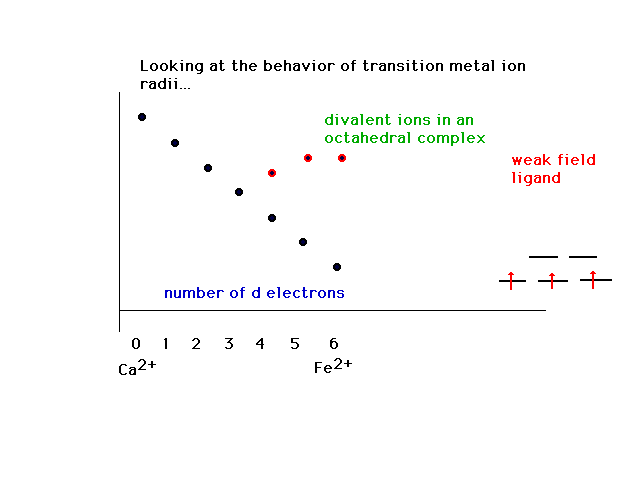
| The counterpart to the above is the change in the
trend when weak field ligands are considered. |
 |
| Changing a transition metal ion's oxidation state has
some understandable consequences. Here, we look at the
shif in color. |
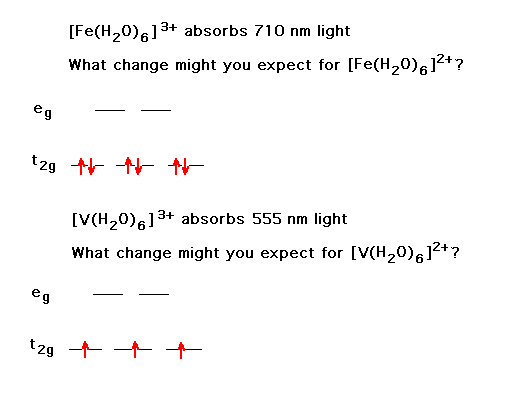 |
| Atomic orbitals in the iron ion. |
 |
| The d-orbital configuration in a strong field
complex. |
 |
| The CO is normally bonded in a linear fashion with
the Fe2+ transition metal ion. |
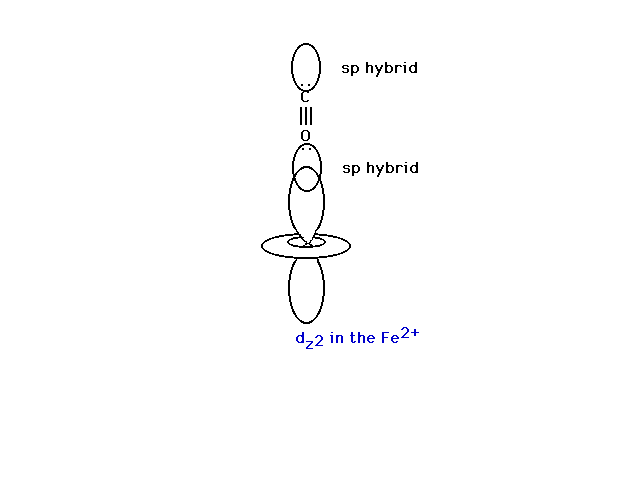 |
| The oxygen molecule is bonded through a bent,
non-linear bond with the Fe2+. |
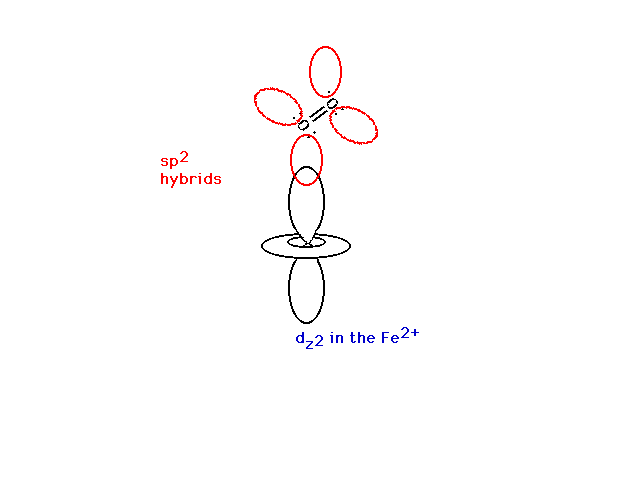 |
| A reminder of the electron states available to
valence electrons in CO. Note the empty pi antibonding
orbitals (whose shapes you should remember). |
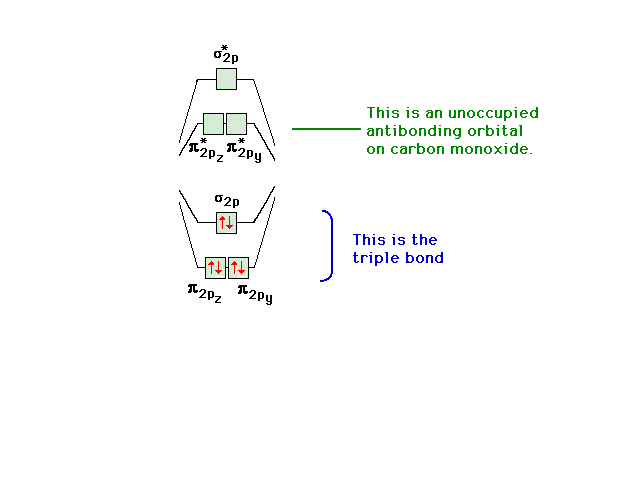 |
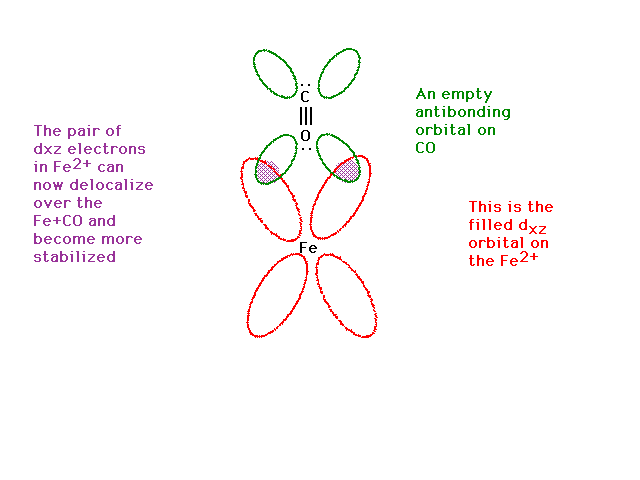
| The dxz is present too. It contains a pair
of electrons (from the iron). |
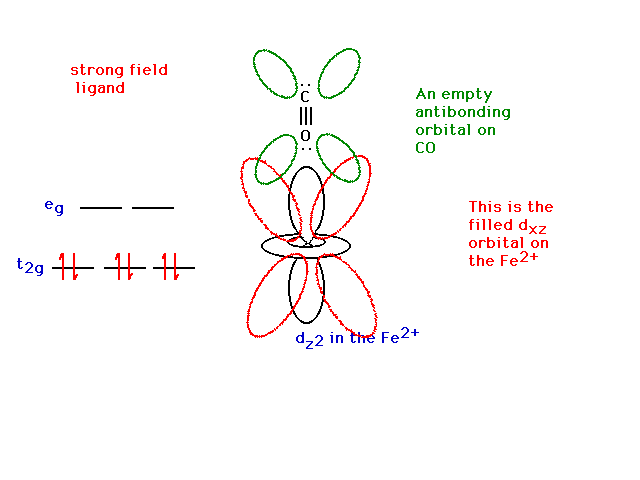 |
| The dxz orbital from the iron and the pi
antibonding orbital from the CO overlap effectively and
give rise to a delocalized orbital involving Fe, C and O.
Yes, you're feeding electrons into the carbon-oxygen bond
antibonding state which would weaken the CO bond, but you
are also strengthening the Fe ligand bond by producing
some pi bond character there. Thus, CO bonds very
strongly to iron (and CO is a strong field ligand). |
 |
| Where have we gotten in our discussion of bonding to
iron in heme? CO is expected to completely tie up all the
iron sites in heme because of the stronger bond than
produced with oxygen binding there. Why are we alive? |
 |
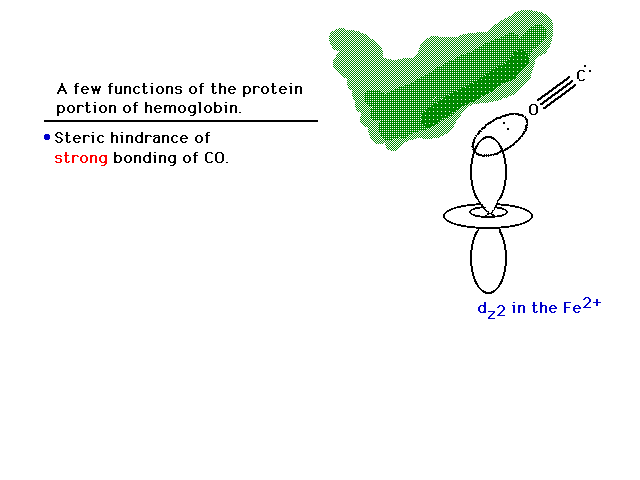
| One explanation is that when the heme is in its
protein, part of the hemoglobin structure forms a canopy
over the oxygen binding site that geometrically prevents
carbon monoxide from binding in a linear fashion. The pi
bond character is eliminated and even the remaining sigma
bond is not too effective, thus allowing oxygen and
carbon monoxide to compete for iron on a more level
playing field. |
 |
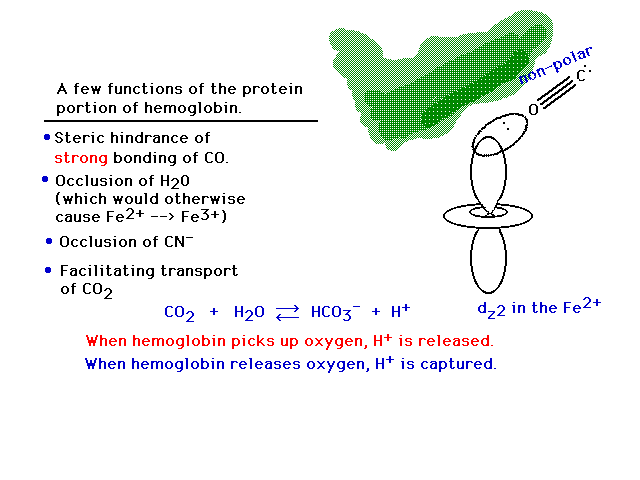
| The side chains influence how the protein affects
chemistry. Non-polar side groups at the entry to the
Fe-heme keep polar species (water, cyanide) away. Oxygen
is non-polar. Carbon monoxide, somewhat surprisingly, has
only 2% ionic character and is therefore basically also
non-polar. |
 |
| The capture and release of H+ by
hemoglobin when oxygen is released to "burn"
glucose or picked up from the air in lungs, respectively,
facilitates the release of CO2 from dissolved
bicarbonate and removal of CO2 waste into
blood plasma at the glucose oxidation sites. |
 |